Effects of Lupenone, Lupeol, and Taraxerol Derived from Adenophora triphylla on the Gene Expression and Production of Airway MUC5AC Mucin
Article information
Abstract
Background
Adenophora triphylla var. japonica is empirically used for controlling airway inflammatory diseases in folk medicine. We evaluated the gene expression and production of mucin from airway epithelial cells in response to lupenone, lupeol and taraxerol derived from Adenophora triphylla var. japonica.
Methods
Confluent NCI-H292 cells were pretreated with lupenone, lupeol or taraxerol for 30 minutes and then stimulated with tumor necrosis factor α (TNF-α) for 24 hours. The MUC5AC mucin gene expression and production were measured by reverse transcription-polymerase chain reaction and enzyme-linked immunosorbent assay, respectively. Additionally, we examined whether lupenone, lupeol or taraxerol affects MUC5AC mucin production induced by epidermal growth factor (EGF) and phorbol 12-myristate 13-acetate (PMA), the other 2 stimulators of airway mucin production.
Results
Lupenone, lupeol, and taraxerol inhibited the gene expression and production of MUC5AC mucin induced by TNF-α from NCI-H292 cells, respectively. The 3 compounds inhibited the EGF or PMA-induced production of MUC5AC mucin in NCI-H292 cells.
Conclusion
These results indicated that lupenone, lupeol and taraxerol derived from Adenophora triphylla var. japonica regulates the production and gene expression of mucin, by directly acting on airway epithelial cells. In addition, the results partly explain the mechanism of of Adenophora triphylla var. japonica as a traditional remedy for diverse inflammatory pulmonary diseases.
Introduction
Mucus in respiratory system plays a pivotal role in defense against invading pathogenic microorganisms, noxious chemicals and particles. The protective function of airway mucus is attributed to the viscoelasticity of mucins. However, any abnormality in the quality or quantity of mucins not only cause altered airway physiology but may also impair host defenses often leading to severe airway pathology as exemplified in chronic bronchitis, cystic fibrosis, asthma, and bronchiectasis1. Therefore, we suggest it is valuable to find the possible activity of controlling (inhibiting) the excessive mucin secretion (production) by various medicinal plants. We have tried to investigate the possible activities of some natural products on mucin secretion from airway epithelial cells. As a result of our trial, we previously reported that several natural products affected mucin secretion and/or production from airway epithelial cells234. According to traditional oriental medicine, Adenophora triphylla var. japonica has been utilised for controlling airway inflammatory diseases5. Also, lupenone, lupeol and taraxerol-its components-were reported to have diverse biological effects including antioxidative and anti-inflammatory effects6789. However, to the best of our knowledge, there are no reports about the potential effects of lupenone, lupeol, and taraxerol on the gene expression and production of mucin from airway epithelial cells. Among the twenty one or more MUC genes coding human mucins reported up to now, MUC5AC was mainly expressed in goblet cells in the airway surface epithelium110. Therefore, we examined the effect of lupenone, lupeol, or taraxerol on tumor necrosis factor α (TNF-α)-induced MUC5AC mucin gene expression and production from NCI-H292 cells, a human pulmonary mucoepidermoid cell line, which are frequently used for the purpose of elucidating intracellular signaling pathways involved in airway mucin production and gene expression111213.
Materials and Methods
1. Materials
All the chemicals and reagents used in this experiment were purchased from Sigma (St. Louis, MO, USA) unless otherwise specified. Lupenone (purity, 98.0%), lupeol (purity, 98.0%), and taraxerol (purity, 98.0%) (Figure 1) were isolated, purified and identified by analytical chemists in the Laboratory of Natural Product Science, Division of Bioscience, Dongguk University (Gyeongju, Korea). Briefly, roots of Adenophora triphylla var. japonica (cultivated in Jeongsun-gun, Gangwon-do, Korea) were provided and authenticated by Professor Je-Hyun Lee (College of Oriental Medicine, Dongguk University, Korea). Air-dried and chopped roots (300 g) were extracted with each of distilled water and 70% ethanol twice at 95℃ for 3 hours. The combined extracts were filtered and concentrated under reduced pressure to give 31.1 g (10.7%) and 38.4 g (12.8%) of crude extracts, respectively. In order to isolate its active constituents the ethanol extract was suspended in distilled water, then consecutively partitioned with organic solvents to give the corresponding hexane (3.6 g), dichloromethane (0.23 g), ethylacetate (0.09 g), and n-butanol (1.4 g) fractions. The main hexane fraction was subjected to column chromatographic separation on a silica gel column using a stepwise elution with dichloromethane, dichloromethane-MeOH 50:1, and dichloromethane-MeOH 1:1 to yield fractions F1-F12. Fraction F3 was recrystallized with dichloromethane-EtOH (two-phase system) 1:1 to give taraxerol (10.8 mg, 0.3%). Fraction F7 was further chromatographed with dichloromethane to afford five subfractions SF1-SF5. Among the subfractions, SF5 was separated by silica gel column chromatography using dichloromethane and MeOH (10:1 to 1:1 gradient) to furnish lupenone (136.8 mg, 3.8%) and lupeol (28.8 mg, 0.8%). The isolated compounds were identified by comparison of their spectral data with literature values141516.
Chemical structure of lupenone (A), lupeol (B), and taraxerol (C).
2. NCI-H292 cell culture
NCI-H292 cells, a human pulmonary mucoepidermoid carcinoma cell line, were purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA) and cultured in RPMI 1640 supplemented with 10% fetal bovine serum (FBS) in the presence of penicillin (100 units/mL), streptomycin (100 µg/mL), and HEPES (25 mM) at 37℃ in a humidified, 5% CO2/95% air, water-jacketed incubator. For serum deprivation, confluent cells were washed twice with phosphate-buffered saline (PBS) and recultured in RPMI 1640 with 0.2% FBS for 24 hours.
3. Treatment of cells with lupenone, lupeol, or taraxerol
After 24 hours of serum deprivation, cells were pretreated with varying concentrations of lupenone, lupeol, or taraxerol for 30 minutes and treated with TNF-α (0.2 nM), epidermal growth factor (EGF; 25 ng/mL), or phorbol 12-myristate 13-acetate (PMA; 10 ng/mL) for 24 hours in serum-free RPMI 1640. Lupenone, lupeol, and taraxerol were dissolved in dimethylsulfoxide and treated in culture medium (final concentrations of dimethylsulfoxide were 0.5%). The final pH values of these solutions were between 7.0 and 7.4. Culture medium and 0.5% dimethylsulfoxide did not affect mucin gene expression and production from NCI-H292 cells. After 24 hours, cells were lysed with buffer solution containing 20 mM Tris, 0.5% NP-40, 250 mM NaCl, 3 mM EDTA, 3 mM EGTA, and protease inhibitor cocktail (Roche Diagnostics, Indianapolis, IN, USA) and collected to measure the production of MUC5AC protein (in 24-well culture plate). The total RNA was extracted for measuring the expression of MUC5AC gene (in 6-well culture plate) by using reverse transcription-polymerase chain reaction (RT-PCR).
4. Total RNA isolation and RT-PCR
Total RNA was isolated by using Easy-BLUE Extraction Kit (Intron Biotechnology, Inc., Seongnam, Korea) and reverse transcribed by using AccuPower RT Premix (Bioneer Co., Daejeon, Korea) according to the manufacturer's instructions. Two micrograms of total RNA was primed with 1 µg of oligo(dT) in a final volume of 50 µL (RT reaction). Two microliters of RT reaction product was polymerase chain reaction (PCR) amplified in a 25 µL by using Thermorprime Plus DNA Polymerase (ABgene, Rochester, NY, USA). Primers for MUC5AC were (forward) 5'-TGA TCA TCC AGC AGG GCT-3' and (reverse) 5'-CCG AGC TCA GAG GAC ATA TGG G-3'. As quantitative controls, primers for Rig/S15 rRNA, which encodes a small ribosomal subunit protein, a housekeeping gene that was constitutively expressed, were used. Primers for Rig/S15 were (forward) 5'-TTC CGC AAG TTC ACC TAC C-3' and (reverse) 5'-CGG GCC GGC CAT GCT TTA CG-3'. The PCR mixture was denatured at 94℃ for 2 minutes followed by 40 cycles at 94℃ for 30 seconds, 60℃ for 30 seconds, and 72℃ for 45 seconds. After PCR, 5 µL of PCR products were subjected to 1% agarose gel electrophoresis and visualized with ethidium bromide under a transilluminator.
5. MUC5AC mucin analysis
MUC5AC airway mucin production was measured by enzyme-linked immunosorbent assay. Cell lysates were prepared with PBS at 1:10 dilution, and 100 µL of each sample was incubated at 42℃ in a 96-well plate, until dry. Plates were washed three times with PBS and blocked with 2% bovine serum albumin (fraction V) for 1 hour at room temperature. Plates were again washed three times with PBS and then incubated with 100 µL of 45M1, a mouse monoclonal MUC5AC antibody (1:200, NeoMarkers, Fremont, CA, USA), which was diluted with PBS containing 0.05% Tween 20 and dispensed into each well. After 1 hour, the wells were washed three times with PBS, and 100 µL of horseradish peroxidase-goat anti-mouse IgG conjugate (1:3,000) was dispensed into each well. After 1 hour, plates were washed three times with PBS. Color reaction was developed with 3,3',5,5'-tetramethylbenzidine peroxide solution and stopped with 1 N H2SO4. Absorbance was read at 450 nm.
6. Statistics
Means of individual group were converted to percent control and expressed as mean±SEM. The difference between groups was assessed using one-way ANOVA and Holm-Sidak test as a post-hoc test. p<0.05 was considered as significantly different.
Results
1. Effect of lupenone, lupeol, or taraxerol on TNF-α-induced MUC5AC gene expression from NCI-H292 cells
As can be seen in Figure 2, MUC5AC gene expression induced by TNF-α from NCI-H292 cells was inhibited by pretreatment with lupenone, lupeol, and taraxerol, respectively (Figure 2). Cytotoxicity was checked by lactate dehydrogenase assay and there was no remarkable cytotoxic effect of lupenone, lupeol, or taraxerol, at the treatment concentrations (data not shown).
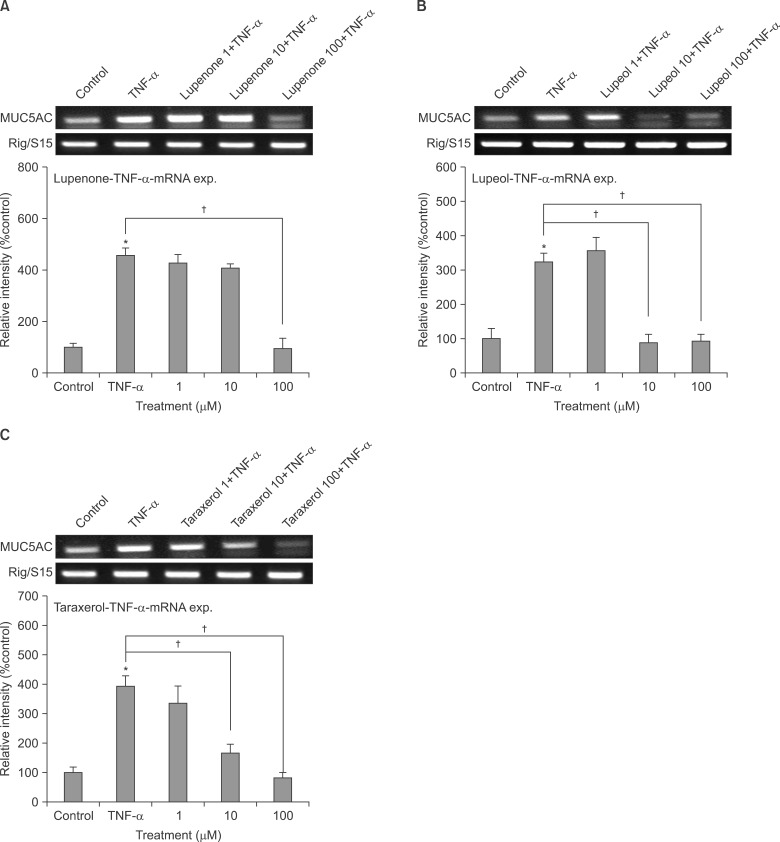
Effect of lupenone (A), lupeol (B), or taraxerol (C) on tumor necrosis factor α (TNF-α)-induced MUC5AC gene expression in NCI-H292 cells. NCI-H292 cells were pretreated with varying concentrations of lupenone, lupeol, or taraxerol for 30 minutes and then stimulated with TNF-α (0.2 nM) for 24 hours. MUC5AC gene expression was measured by reverse transcription-polymerase chain reaction. Three independent experiments were performed and the representative images were shown. Polymerase chain reaction images were quantified using densitometry and the graphic data were also shown. Each bar represents a mean±SEM of 3 densitograms in comparison with that of control set at 100% (A-C). *Significantly different from control (p<0.05). †Significantly different from TNF-α alone (p<0.05).
2. Effect of lupenone on TNF-α-induced MUC5AC mucin production from NCI-H292 cells
Lupenone inhibited TNF-α-induced MUC5AC mucin production. The amounts of MUC5AC mucin in the cells of lupenone-treated cultures were 100±6%, 543±29%, 362±14%, 324±7%, and 114±7% for control, 0.2 nM of TNF-α alone, TNF-α plus lupenone 10-6 M, TNF-α plus lupenone 10-5 M, and TNF-α plus lupenone 10-4 M, respectively (Figure 3A).
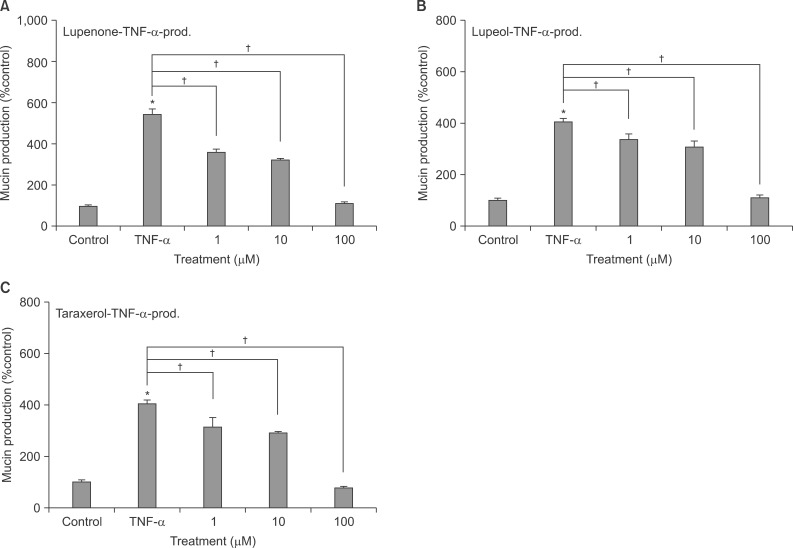
Effect of lupenone (A), lupeol (B), or taraxerol (C) on tumor necrosis factor α (TNF-α)-induced MUC5AC mucin production in NCI-H292 cells. NCI-H292 cells were pretreated with varying concentrations of lupenone, lupeol or taraxerol for 30 minutes and then stimulated with TNF-α (0.2 nM, 10 ng/mL) for 24 hours. Cell lysates were collected for measurement of MUC5AC mucin production by enzyme-linked immunosorbent assay. Each bar represents a mean±SEM of three culture wells in comparison with that of the control set at 100% (A-C). Three independent experiments were performed and the representative data were shown. *Significantly different from control (p<0.05). †Significantly different from TNF-α alone (p<0.05).
3. Effect of lupeol on TNF-α-induced MUC5AC mucin production from NCI-H292 cells
Lupeol also inhibited TNF-α-induced MUC5AC mucin production. The amounts of MUC5AC mucin in the cells of lupeol-treated cultures were 100±10%, 405±13%, 337±21%, 307±24%, and 110±11% for control, 0.2 nM of TNF-α alone, TNF-α plus lupeol 10-6 M, TNF-α plus lupeol 10-5 M, and TNF-α plus lupeol 10-4 M, respectively (Figure 3B).
4. Effect of taraxerol on TNF-α-induced MUC5AC mucin production from NCI-H292 cells
Taraxerol also inhibited TNF-α-induced MUC5AC mucin production. The amounts of MUC5AC mucin in the cells of taraxerol-treated cultures were 100±10%, 405±13%, 315±36%, 290±7%, and 77±7% for control, 0.2 nM of TNF-α alone, TNF-α plus taraxerol 10-6 M, TNF-α plus taraxerol 10-5 M, and TNF-α plus taraxerol 10-4 M, respectively (Figure 3C).
5. Effect of lupenone on EGF-induced MUC5AC mucin production from NCI-H292 cells
Lupenone significantly inhibited EGF-induced MUC5AC production from NCI-H292 cells. The amounts of mucin in the cells of lupenone-treated cultures were 100±13%, 330±14%, 232±2%, 116±8%, and 75±4% for control, 25 ng/mL of EGF alone, EGF plus lupenone 10-6 M, EGF plus lupenone 10-5 M, and EGF plus lupenone 10-4 M, respectively (Figure 4A).
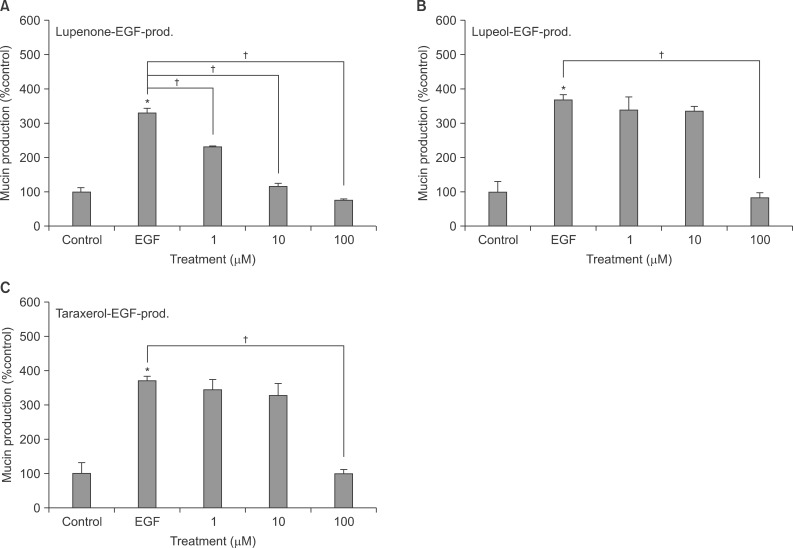
Effect of lupenone (A), lupeol (B), or taraxerol (C) on epidermal growth factor (EGF)-induced MUC5AC mucin production in NCI-H292 cells. NCI-H292 cells were pretreated with varying concentrations of lupenone, lupeol, or taraxerol for 30 minutes and then stimulated with EGF (25 ng/mL) for 24 hours. Cell lysates were collected for measurement of MUC5AC mucin production by enzyme-linked immunosorbent assay. Each bar represents a mean±SEM of three culture wells in comparison with that of control set at 100% (A-C). Three independent experiments were performed and the representative data were shown. *Significantly different from control (p<0.05). †Significantly different from EGF alone (p<0.05).
6. Effect of lupeol on EGF-induced MUC5AC mucin production from NCI-H292 cells
Lupeol significantly inhibited EGF-induced MUC5AC production from NCI-H292 cells. The amounts of mucin in the cells of lupeol-treated cultures were 100±31%, 369±15%, 340±38%, 336±13%, and 79±18% for control, 25 ng/mL of EGF alone, EGF plus lupeol 10-6 M, EGF plus lupeol 10-5 M, and EGF plus lupeol 10-4 M, respectively (Figure 4B).
7. Effect of taraxerol on EGF-induced MUC5AC mucin production from NCI-H292 cells
Taraxerol significantly inhibited EGF-induced MUC5AC production from NCI-H292 cells. The amounts of mucin in the cells of taraxerol-treated cultures were 100±31%, 369±15%, 344±29%, 327±36%, and 100±11% for control, 25 ng/mL of EGF alone, EGF plus taraxerol 10-6 M, EGF plus taraxerol 10-5 M, and EGF plus taraxerol 10-4 M, respectively (Figure 4C).
8. Effect of lupenone on PMA-induced MUC5AC mucin production from NCI-H292 cells
Lupenone significantly inhibited PMA-induced MUC5AC production from NCI-H292 cells. The amounts of mucin in the cells of lupenone-treated cultures were 100±14%, 247±2%, 261±1%, 147±8%, and 55±3% for control, 10 ng/mL of PMA alone, PMA plus lupenone 10-6 M, PMA plus lupenone 10-5 M, and PMA plus lupenone 10-4 M, respectively (Figure 5A).

Effect of lupenone (A), lupeol (B), or taraxerol (C) on phorbol 12-myristate 13-acetate (PMA)-induced MUC5AC mucin production in NCI-H292 cells. NCI-H292 cells were pretreated with varying concentrations of lupenone, lupeol, or taraxerol for 30 minutes and then stimulated with PMA (10 ng/mL) for 24 hours. Cell lysates were collected for measurement of MUC5AC mucin production by enzyme-linked immunosorbent assay. Each bar represents a mean±SEM of three culture wells in comparison with that of control set at 100% (A-C). Three independent experiments were performed and the representative data were shown. *Significantly different from control (p<0.05). †Significantly different from PMA alone (p<0.05).
9. Effect of lupeol on PMA-induced MUC5AC mucin production from NCI-H292 cells
Lupeol significantly inhibited PMA-induced MUC5AC production from NCI-H292 cells. The amounts of mucin in the cells of lupeol-treated cultures were 100±6%, 273±6%, 262±14%, 205±7%, and 123±15% for control, 10 ng/mL of PMA alone, PMA plus lupeol 10-6 M, PMA plus lupeol 10-5 M, and PMA plus lupeol 10-4 M, respectively (Figure 5B).
10. Effect of taraxerol on PMA-induced MUC5AC mucin production from NCI-H292 cells
Taraxerol significantly inhibited PMA-induced MUC5AC production from NCI-H292 cells. The amounts of mucin in the cells of taraxerol-treated cultures were 100±5%, 378±24%, 426±48%, 308±14%, and 121±7% for control, 10 ng/mL of PMA alone, PMA plus taraxerol 10-6 M, PMA plus taraxerol 10-5 M, and PMA plus taraxerol 10-4 M, respectively (Figure 5C).
Discussion
TNF-α has been reported to stimulate the secretion and gene expression of airway mucin131718. TNF-α converting enzyme provoked MUC5AC mucin expression in cultured human airway epithelial cells13 and TNF-α-induced MUC5AC gene expression in normal human airway epithelial cells18. TNF-α level in sputum was reported to be increased, with further increases during exacerbation of pulmonary diseases1920. It also induced mucin secretion from guinea pig tracheal epithelial cells17. Based upon these reports, we investigated whether lupenone, lupeol, or taraxerol affects TNF-α-induced MUC5AC mucin gene expression and production from NCI-H292 cells. As shown in results, MUC5AC mucin gene expression induced by TNF-α from NCI-H292 cells was inhibited by pretreatment with lupenone, lupeol, and taraxerol, respectively (Figure 2). At the same time, lupenone, lupeol or taraxerol suppressed TNF-α-induced production of MUC5AC mucin protein (Figure 3). These results suggest that lupenone, lupeol, or taraxerol can regulate the gene expression and production of MUC5AC mucin induced by TNF-α, by directly acting on airway epithelial cells.
Next, we tried to investigate whether lupenone, lupeol, or taraxerol affects MUC5AC production induced by EGF or PMA, the other well-known stimulator of mucin production from airway epithelial cells. EGF has been reported to regulate MUC5AC gene expression in the lung. MUC5AC mRNA expression was reported to increase after ligand binding to the EGF receptor and activation of the mitogen-activated protein kinase (MAPK) cascade1219. On the other hand, PMA was reported to stimulate the endogenous activator of protein kinase C (PKC), diacylglycerol21 and to be an inflammatory stimulant that can control a gene transcription22, cell growth and differentiation23. PMA also can induce MUC5AC gene expression in NCI-H292 cells24. PMA activates a type of PKC isoforms. This activates matrix metalloproteinases, which cleave pro-EGF receptor ligands from the cell surface to become mature EGF receptor ligands. These ligands bind to the EGF receptor, provoking the phosphorylation of its intracellular tyrosine kinase. This leads to activation of MEK leading to ERK activation. Following is the activation of the transcription factor, Sp1, and binding of the factor to specific sites with the MUC5AC gene promoter. Finally, the promoter is activated and produced the gene transcription and translation to MUC5AC mucin protein22. As shown in results, EGF-induced production of MUC5AC mucin protein was suppressed by pretreatment of lupenone, lupeol, and taraxerol, respectively (Figure 4). Also, PMA-induced production of MUC5AC mucin protein was inhibited by pretreatment of lupenone, lupeol, and taraxerol, respectively (Figure 5).
The underlying mechanisms of action of lupenone, lupeol and taraxerol on MUC5AC mucin gene expression and production are not clear at present, although we are investigating whether lupenone, lupeol and taraxerol act as potential regulators of nuclear factor kB signaling pathway and/or the MAPK cascade after ligand binding to the TNF or EGF receptor, in mucin-producing NCI-H292 cells.
Taken together, the inhibitory action of lupenone, lupeol or taraxerol on airway mucin production and gene expression might explain, at least in part, the traditional use of Adenophora triphylla var. japonica as an anti-inflammatory mucoregulator for pulmonary inflammatory diseases, in folk medicine. We suggest it is valuable to find the natural products that have specific inhibitory effects on mucin production and gene expression-in view of both basic and clinical sciences-and the result from this study suggests a possibility of using lupenone, lupeol or taraxerol as a new efficacious mucoregulator for pulmonary diseases, although further studies are essentially required.
Acknowledgements
This research was supported by a grant (12172KFDA989) from Korea Food & Drug Administration in 2012.
Notes
Conflicts of Interest: No potential conflict of interest relevant to this article was reported.