Introduction
Skeletal muscle, the most abundant tissue in the human body, is essential for the maintenance of body composition and physical activity. Muscle wasting, a phenomenon that occurs when there is an imbalance between the rates of protein synthesis and degradation, is commonly seen in patients with sepsis and other catabolic human diseases1. Muscle wasting may lead to weakness and fatigue that negatively affects ambulation and increases the risk for thromboembolic complications. The involvement of respiratory muscles increases the risk of pulmonary complications and may necessitate or prolong the need for ventilatory support2.
Sepsis, a systemic inflammatory response that results from an infection due to the presence of microorganisms or their toxins in the host tissue, is frequently associated with organ system dysfunction3. Muscle catabolism in sepsis is caused by inhibition of protein synthesis in conjunction with increased protein degradation1,4. In addition to the physically detrimental aspects of muscle wasting, persistent wasting during sepsis leads to muscle dysfunction and a prolonged recovery time. Sepsis is typically characterized by a cytokine storm, which represents a pro-inflammatory cytokine/anti-inflammatory cytokine imbalance5. Markedly elevated levels of various cytokines, like tumor necrosis factor α (TNF-α) and interleukin-6, during sepsis may influence muscle wasting and protein degradation6. It has been shown that treatment with TNF-α stimulates time- and concentration-dependent reductions in the total protein content and loss of fast-type myosin heavy chain (MyHC) content in cultured myotubule7.
Recently, interleukin-15 (IL-15), a cytokine, has been implicated in skeletal myogenesis. IL-15 (14~15 kDa) is a 4-helix cytokine known to stimulate T-cell proliferation and enhance natural killer cell activity8. Tissue culture studies have demonstrated that IL-15 induces the accumulation of MyHCs in differentiated muscle cells9, thereby strengthening evidence that IL-15 is a novel anabolic agent that increases skeletal muscle mass10 and in this way influences muscle growth. Similar studies have also shown that IL-15 stimulates myogenic differentiation independently of insulin-like growth factors (IGFs)11. However, to date, the role of IL-15 in sepsis-induced muscle wasting remains unclear. In view of the above, the objective of the present study was to investigate the role of IL-15 in sepsis-induced skeletal muscle atrophy and proteolysis in vitro and in vivo.
Materials and Methods
1. Cell culture
A mouse skeletal muscle cell line (C2C12) was obtained from the American Type Culture Collection (Rockville, MD, USA) and employed for in vitro studies. C2C12 cells were passaged in high-glucose Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin, 100 mg/mL streptomycin, 25 ng/mL fungizone, 110 mg/mL sodium pyruvate, and 2 mM L-glutamine in a humidified atmosphere of 5% CO2 and 95% air, at 37℃. For in vitro studies, cells were seeded at 5×104 cells/cm2 in 10% FBS/DMEM until they reached 70~80% confluence.
2. In vitro experiments
IL-15 antibodies and mouse recombinant IL-15 were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). C2C12s were stimulated in separate treatments with either dexamethasone (10 or 100 nM), lipopolysaccharide (LPS; 10 ng/mL, 20 ng/mL, or 1µg/mL), or TNF-α (10µg/mL) to induce muscle proteolysis (all from Sigma-Aldrich, St. Louis, MO, USA). To evaluate the effect of IL-15 on muscle proteolysis, Cells were stimulated with either LPS or dexamethasone in the presence and absence of IL-15, and sampled at different time intervals (24, 48, or 72 hours) after stimulation.
3. Measurement of proteolysis
Cathepsin L activity was determined as a measure of muscle proteolysis in the culture supernatant from co-stimulated C2C12 cells and lysates of muscle tissues. Cathepsin L activity was determined by measuring the degradation of the fluorogenic cathepsin L-specific substrate in lysates from stimulated C2C12 cells and muscle tissues. Cathepsin L activity was analyzed using an activity assay kit (Biovision, Inc., Mountain View, CA, USA), according to the manufacturer's instructions.
4. In vivo experiment
For in vivo studies, BALB/C mice (67 weeks of age) were acquired from Koatech (Cheonan, Korea). Sepsis was induced either by cecal ligation and puncture (CLP) or intraperitoneal administration of LPS (10 mg/kg i.p.; Sigma-Aldrich)12. The murine CLP model that reproduces many of the clinical features of sepsis in patients was used to induce intra-abdominal sepsis. Briefly, after externalization of the cecum, perforation was made on the antimesenteric border of the cecum with an 18-gauge needle and the cecum ligated 2-cm distal to the ileocecal valve. Postoperatively, mice were resuscitated with 0.5 mL of normal saline intraperitoneally.
IL-15 (10µg/kg) was intraperitoneally administered 6 hours before the induction of sepsis with CLP, and limb muscles were sampled 24 hours after induction of sepsis. The cathepsin L activity was also determined in the lysate of the recovered limb muscle samples and compared with that of the control. Samples of limb muscles, that is, the gastrocnemius (GN), extensor digitorum longus (EDL), and tibialis anterior (TA) were obtained 16 hours after induction of sepsis. Subsequently, expression of the skeletal muscle atrophy-related genes, atrogin-1 and muscle-specific ring finger protein 1 (MuRF1)13, were analyzed in limb muscle samples by using reverse transcriptase polymerase chain reaction (RT-PCR) and western blot analyses, and compared with that of the control. The number of animals for the in vivo experiment was 3 control mice, 5 in the CLP or LPS sepsis group, and 5 in CLP with IL-15 administration group. All groups were treated concurrently.
5. RT-PCR and Western blot
For semi-quantitative RT-PCR, 1µg of the isolated total RNA was reverse transcribed using the iScript cDNA synthesis kit (Bio-Rad, Hercules, CA, USA) according to the manufacturer's instruction. Targeted genes were amplified by PCR with the following set of primers (sense and antisense, respectively): 5'-AACATGTGGGTGTATCGG-3', 5'-TCTTGAGGGGAAAGTGAG-3', 464 bp for the Atrogin-1; 5'-GTTAAACCAGAGGTTCGAG-3', 5'-ATGGTTCGCAACATTTCGG-3', 342 bp for the MuRF1; 5'-GTCTTCATTTTGGGCTGTTTCAGT-3', 5'-CCTCACATTCTTTGCATCCAGATTCT-3', 315 bp for the IL-15 ; 5'-AATGCATCCGCACCACCAA-3', 5'-GTAGCCATATTCATTGTCAT-3', 515 bp for the glyceraldehyde-3-phosphate dehydrogenase. Each amplification cycle consisted of denaturation at 94℃ for 30 seconds, annealing at 58℃ for 1 minute, and extension at 72℃ for 1 minute. PCR reactions were performed for 30 amplification cycles. Samples were electrophoresed on a 1.2% agarose gel in Tris-acetic acid-EDTA buffer and stained with ethidium bromide. The bands were viewed under UV light and The resulting gel image files analyzed by densitometric analysis using the PD-Quest software (Bio-Rad).
For Western blot analysis, Tissue samples were homogenized on ice in RIPA buffer containing a protease inhibitor cocktail (Sigma-Aldrich). The homogenate was sonicated and centrifuged at 12,000 ×g for 10 minutes at 4℃ to remove insoluble debris. The protein concentration was measured using BCA protein assay kit (Pierce, Rockford, IL, USA). Equal protein amounts of limb muscles extracts were resolved on a 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis. The separated proteins were electrophoretically transferred onto a nitrocellulose membrane using the semidry technique. The membrane was blocked for 1 hour in TBST solution (Tris-buffed saline, 20 mM Tris-HCl, pH 7.6, 137 mM NaCl, and 0.1% Tween 20) containing 5% skim milk. After blocking, the membranes were washed in TBST twice for 10 minutes each and incubated with anti-IL-15 antibody overnight at 4℃. After additional washes with TBST, the membrane was subsequently incubated with the appropriate horseradish peroxidase-conjugated secondary antibody (1:5,000) for 1 hour. After final washes, proteins were visualized by the enhanced chemiluminescence method (Pierce). β-actin was used as a loading control in the stripped blot.
Results
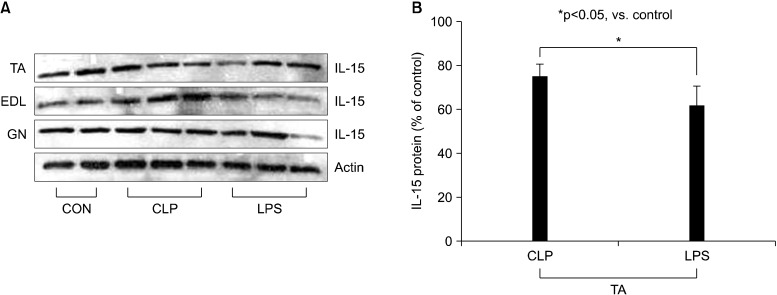
In the present study, in the sepsis model described, both mRNA and protein expression of IL-15 was significantly lower in the limb muscle samples obtained from septic mice than in the controls. IL-15 mRNA expression was measured with RT-PCR in the TA and EDL limb muscle samples of septic mice induced by CLP and intraperitoneal injection of LPS (10 mg/kg i.p.) at 16 hours after sepsis induction. When IL-15 mRNA expression in these mice was compared to the control, using densitometry, IL-15 mRNA expression was significantly lower in both TA and EDL muscle samples of septic mice than in the control mice (p<0.05) (Figure 1). IL-15 protein expression was also measured in the TA, EDL, and GN limb muscle samples of septic mice by western blot analysis at 16 hours after sepsis induction. When IL-15 protein levels were compared to those of the control, using densitometry, IL-15 protein expression was significantly lower only in TA muscle samples of septic mice when compared to that of the control (p<0.05) (Figure 2). The mRNA expression of muscle atrophy-related genes, atrogin-1 and MuRF1, was analyzed by RT-PCR in limb muscle samples derived from TA and EDL samples of septic mice. We observed that the mRNA expression of both atrogin-1 and MuRF1 were significantly higher in TA and EDL muscle samples of septic mice than in the control (p<0.05) (Figure 3). This indicated up-regulation of expression of muscle atrophy-related genes in sepsis.
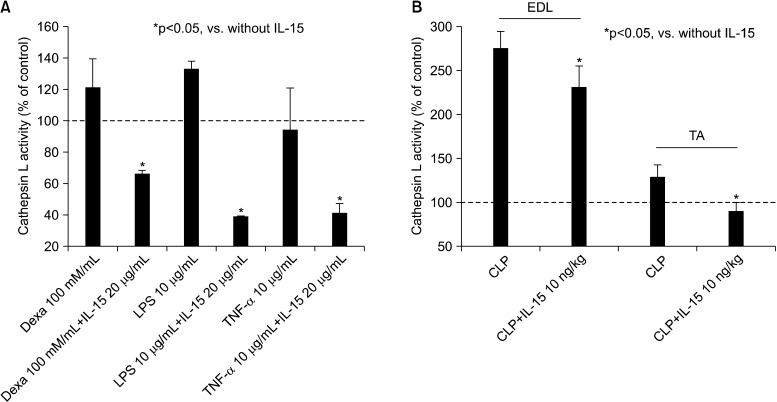
When cultured mouse myoblast cells (C2C12) were stimulated in separate treatments with either dexamethasone (10 and 100 nM) or LPS (10, 20, or 1µg/mL) or TNF-α (10µg/mL), we observed that the rate of muscle cathepsin L activity increased over time, indicating the occurrence of muscle proteolysis. However, when the cells were co-stimulated in the presence of IL-15 along with either LPS, dexamethasone, or TNF-α, a marked reduction in the level of cathepsin L activity was observed, in comparison to the enzymatic activity observed when either LPS, dexamethasone, or TNF-α was used alone (p<0.05) (Figure 4). In addition, we monitored cathepsin L enzyme activity in TA and EDL muscle lysates of mice pretreated with IL-15 (10µg/kg), prior to the induction of sepsis by CLP. A significant decrease was observed in cathepsin L activity in TA and EDL muscle lysates of mice pretreated with IL-15, when compared to those obtained from mice that had not been pretreated (p<0.05) (Figure 4).
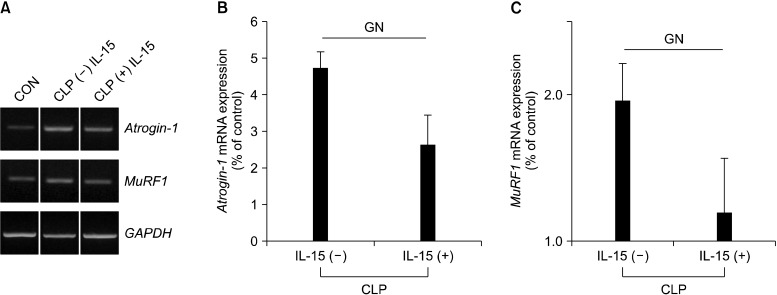
Similarly, mRNA expression of atrogin-1 and MuRF1 was evaluated by RT-PCR in muscle lysates derived from GN limb muscle samples of mice pretreated with IL-15 prior to the induction of sepsis by CLP. Notably, as seen in Figure 5, there was a marked decrease in the mRNA expression levels of both genes in the presence of IL-15 when compared to those obtained from mice that had not been pretreated with IL-15, as well as from control mice (p<0.05) (Figure 5).
Discussion
To the best of our knowledge, this is the first study to demonstrate the role of IL-15 in sepsis-induced muscle wasting. The expression of IL-15 was found to be significantly lower in limb muscles of septic mice than in the control. Our results are supported by a previous study that demonstrated the anabolic effect of IL-15 in muscle cell culture and showed a decrease in the muscle degradation rate in a cachexia animal model10,14, suggesting that IL-15 is important in muscle growth. Thus, the present study confirms that IL-15 has an anabolic effect on muscle growth and antagonizes muscle degradation and leads to the further hypothesis that a decrease in the expression of IL-15 in the limb muscles of the sepsis animal model may contribute to sepsis-induced muscle wasting.
In our study, the expression levels of IL-15 in the limb muscles were significantly higher than those in the diaphragm of the septic mice (data not shown). Limb muscles are dominated by type II muscle fibers, whereas the diaphragm is usually composed of type I muscle fibers. Our results are in line with those of a previous study that demonstrated that the mRNA levels of IL-15 were comparatively higher in human skeletal muscles dominated by type II fast-twitch muscle fibers than in muscles dominated by type I slow-twitch muscle fibers15. Furthermore, muscle atrophy during sepsis has been shown to be fiber type-specific in skeletal muscles, mostly affecting the fast-twitch type IIa muscle fibers16. Taken together, the high expression levels of IL-15 in limb muscles indicate that IL-15 is usually expressed in type II fast-twitch muscle fibers, and thus plays an important role in sepsis-induced muscle atrophy.
The present study, through in vivo and in vitro experiments, further demonstrated a marked decrease in the activity of cathepsin L enzyme in the presence of IL-15, leading to a decrease in muscle protein degradation. In addition, the expression of atrogin-1 and MuRF1 was found to be decreased in the limb muscles of septic mice. These results indicate that the decrease in muscle proteolysis by IL-15 is associated with a lower expression of atrogin-1 and MuRF1, which are the key elements regulating the ubiquitin-proteasome-mediated muscle protein degradation pathway1,4,17, thus suggesting a plausible role for IL-15 in proteolysis.
Several studies have indicated IL-15 as a therapeutic target for treating muscle-wasting disorders. A previous study demonstrated that C2C12 myotubes transfected with IL-15 showed a marked increase in myofibrillar protein accumulation and inhibited protein degradation in skeletal myogenic cell cultures, in contrast to IGF-I, which only stimulated protein synthesis under culture conditions9. This finding suggested that the protein accumulation effect of IL-15 occurred without stimulation of skeletal myoblast proliferation or differentiation, while IGF-1, being a muscle anabolic factor, acted indirectly by stimulating myoblast proliferation and differentiation11. A similar study using an animal model of hepatoma-inducing cachexia14 demonstrated that IL-15 administration prevents limb muscle wasting via inhibiting protein turnover and thus highlighted its therapeutic effect. Another study, on tumor-bearing rats, showed that IL-15 treatment attenuated muscle wasting in rats with cancer cachexia and further attributed this mechanism to the lower expression levels of the components of the ubiquitin-proteasome pathway18. It has also been shown that 4 weeks of IL-15 treatment improve diaphragm strength, which is in turn associated with increased muscle fiber cross-sectional area and decreased collagen infiltration in Duchenne muscular dystrophy19. Collectively, these results indicate that IL-15 has possible therapeutic potential in the treatment of various muscle wasting disorders.
In conclusion, the present study demonstrated a significant decrease in IL-15 expression in the limb muscles of a sepsis animal model and further showed that muscle proteolysis and atrophy-related gene expression were greatly attenuated in the limb muscles of septic mice in the presence of IL-15. Our findings lend support to the hypothesis that IL-15 plays a critical role in protection against sepsis-induced muscle wasting and may therefore serve as a potential therapeutic target for treating sepsis-induced skeletal muscle wasting and muscle proteolysis.


 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Print
Print Download Citation
Download Citation